Les proies
 Durant la phase d’hivernage des limicoles dans les Pertuis charentais l’acquisition des ressources alimentaires est une nécessité vitale et va amplement conditionner le comportement des oiseaux. Il apparait donc indispensable de connaitre les liens entre les prédateurs et leurs proies et d’estimer le stock de proies disponibles et sa variabilité.
Durant la phase d’hivernage des limicoles dans les Pertuis charentais l’acquisition des ressources alimentaires est une nécessité vitale et va amplement conditionner le comportement des oiseaux. Il apparait donc indispensable de connaitre les liens entre les prédateurs et leurs proies et d’estimer le stock de proies disponibles et sa variabilité.
Après avoir décrit la diversité, les abondances, les phénologies et les distributions des oiseaux limicoles, il convient d’opérer le même travail sur les proies potentielles des principales espèces de limicoles, essentiellement constituées par la macrofaune benthique des vasières intertidales.
Dans les Pertuis Charentais les espèces de proies dominantes sont les mollusques tels que les bivalves (Macoma balthica, Cerastoderma edule, Scrobicularia plana et Abra tenuis) ou les gastéropodes (Hydrobia ulvae) et quelques espèces d’annélides vont constituer le gros de la biomasse en vers telles que Nephtys hombergii et Hedistes diversicolor. D’autres sites tels que des vasières recouvertes par des herbiers à Zostères Zostera noltii vont abriter des communautés différentes avec une raréfaction des mollusques dominants comme Macoma balthica ou Abra tenuis au profit d’autres espèces comme Tapes spp ou Abra alba. Dans le cadre de cette étude, nous avons également pris en compte les abondances et la distribution de la zostère naine Zostera noltii sur l’ensemble des sites.
Les principales proies
Les bivalves
</i>")
</i>")
</i>")
</i> (© ecomare)")
</i>)")
Les gastéropodes
</i>")
")
Les vers
</i>")
</i>")
</i> © Philippe Giraud, biosphoto")
")
Les crustacés
</i>")
")
")
Les plantes
</i>")
En savoir plus ...
La stratégie d’échantillonnage
L’échantillonnage systématique des surfaces abritant la macrofaune benthique doit nous permettre d’identifier les espèces proies plus abondantes à travers le calcul des occurrences et des densités mais aussi leurs disponibilités, leurs profitabilités et leurs accessibilités pour les oiseaux. La comparaison des abondances et des diversités de la macrofaune par site permet de mettre en évidence leurs valeurs trophiques relatives pour les prédateurs. Le même quadrillage de prélèvements a été appliqué sur plusieurs sites des Pertuis à raison de 250 m entre stations. Pour les stations échantillonnées à pied une carotte de sédiments de 0,018 (1/56) m² est prélevée sur une profondeur d’environ 20-25 cm. Dans certaines zones pour des questions pratiques et d’accès, un échantillonnage par bateau a été effectué. En bateau, deux carottes de sédiments de 0,008 m² et 20-25 cm de profondeur ont été réalisées soit une surface totale de 0,016 m². Sur le terrain les carottes sont tamisées sur vide de maille de 1 mm. Les gastéropodes de petites tailles essentiellement représentés par Hydrobia ulvae, sont échantillonnés par carotte de 0,0037 m² pour une profondeur de 4-5 cm et ensuite tamisés sur vide de maille de 500 µm. En bateau, les hydrobies sont échantillonnées uniquement sur la deuxième carotte et tamisées sur vide de maille de 500 µm. Tous les organismes vivants sont triés et conservés au congélateur à -20°C dans des sacs plastiques jusqu’à leur traitement au laboratoire. Les crustacés et vers sont isolés et conservés dans de l’alcool à 70°. Au laboratoire, les organismes sont identifiés jusqu’à l’espèce lorsque cela est possible. Chaque individu est mesuré avec une précision de l’ordre du 10ème de millimètre pour déterminer les gammes de taille des proies disponibles pour les oiseaux. Les masses sèches (DM dry Mass) et les masses sèches sans cendre (Ash Free Dry Mass AFDM) sont établies pour la chair des mollusques de manière à disposer de valeur nutritive moyenne par classe de taille. Les annélides ayant été conservés à l’alcool, les masses sont reconstituées à partir des tailles grâce à des relations allométriques.
Les sédiments
Un échantillon de sédiments d’une surface de 0,0037 m² sur une profondeur d’environ 3 cm est aussi prélevé sur une station sur quatre de façon à décrire la structure morpho-sédimentaire des vasières étudiées. Ces prélèvements couvrent une majorité des secteurs de chaque site et ceci pour différents niveaux hypsométriques.
Les variations spatiales
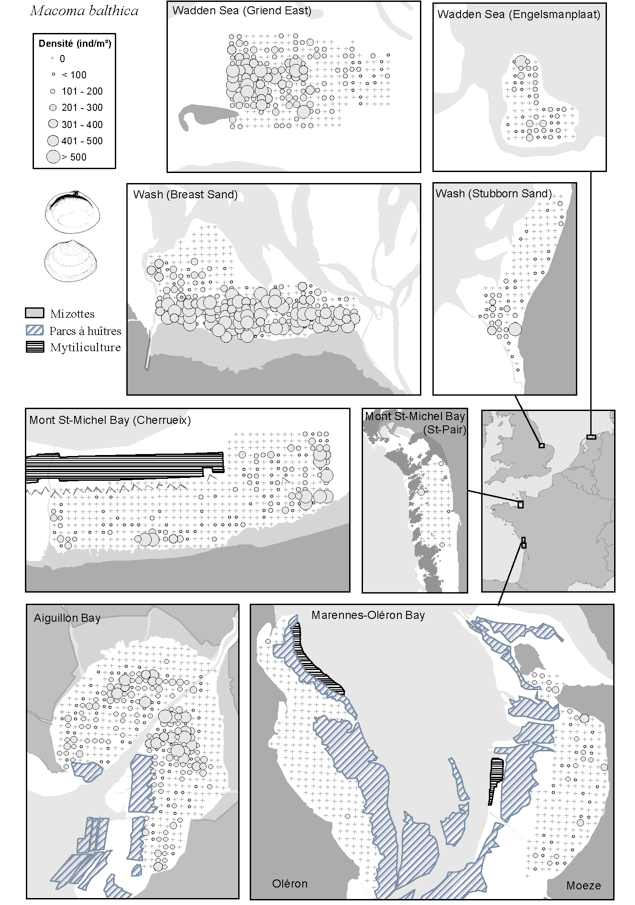
Les données issues de cet échantillonnage ont dans un premier temps été utilisées pour décrire et comparer la distribution spatiale des espèces de mollusques les plus communes sur un gradient latitudinal de 8° (46-54°N) et 900 km de distance (Bocher et al. 2007). Parmi les quinze espèces de bivalves et huit espèces de gastéropodes collectées, nous avons retenu les quatre espèces les plus communes et les plus abondantes de bivalves (Cerastoderma edule, Macoma balthica, Scrobicularia plana and Abra tenuis), qui représentent à elles seules 96% de l’ensemble des bivalves prélevés, et l’hydrobie Hydrobia ulvae qui comprend 99% de l’ensemble des gastéropodes. C. edule et M. balthica sont les bivalves les plus répandus et les plus communs sur les vasières intertidales en Europe avec des densités maximales supérieures à 150 ind./m2 mesurées aux plus hautes latitudes. S. plana et A. tenuis sont plus abondants sur les sites du sud avec une claire (ou nette) préférence pour les vasières à granulométrie très fine. L’hydrobie apparaît communément sur tous les sites et en densités comparables excepté en Baie du Mont Saint-Michel où elle est extrêmement rare. Les tailles moyennes des mollusques varient grandement entre les sites sans gradient apparent du nord au sud. Les patrons de distribution des mollusques sur les cinq vasières intertidales des neufs sous-sites étudiés sont majoritairement spécifiques à chaque site. La distribution des M. balthica est montrée à titre d’exemple parmi les cinq espèces. L’étude a ainsi permis de montrer que la composition des communautés de mollusques possède de plus grandes similarités à l’intérieur des régions situées de parts et d’autres de la Bretagne plutôt qu’entre elles.
Les assemblages
Les assemblages d’espèces sont souvent associés avec la diversité des structures des habitats. Les systèmes sédimentaires ne semblent pas y faire exception. Ainsi, à l’intérieur des systèmes sédimentaires, la diversité des espèces benthiques apparaît être corrélée à la complexité granulométrique (Compton et al. 2008). Le but de cette étude était d’examiner si les diversités des bivalves sont associées à l’hétérogénéité sédimentaire au sein même du système et si la complexité de la granulométrie à l’intérieur de neufs systèmes sédimentaires étudiés diffère selon une situation en milieu tempéré ou tropical. Bien que les stratégies d’histoires de vie des bivalves, comme la sélection de l’habitat après le recrutement, suggèrent que l’hétérogénéité du sédiment est importante pour les espèces de bivalves, la diversité des bivalves et l’hétérogénéité du sédiment ne sont pas associées au travers des différents systèmes. La diversité en bivalves sur un site est beaucoup plus importante dans les complexes avec des sédiments fins.
Du total au « collectable »
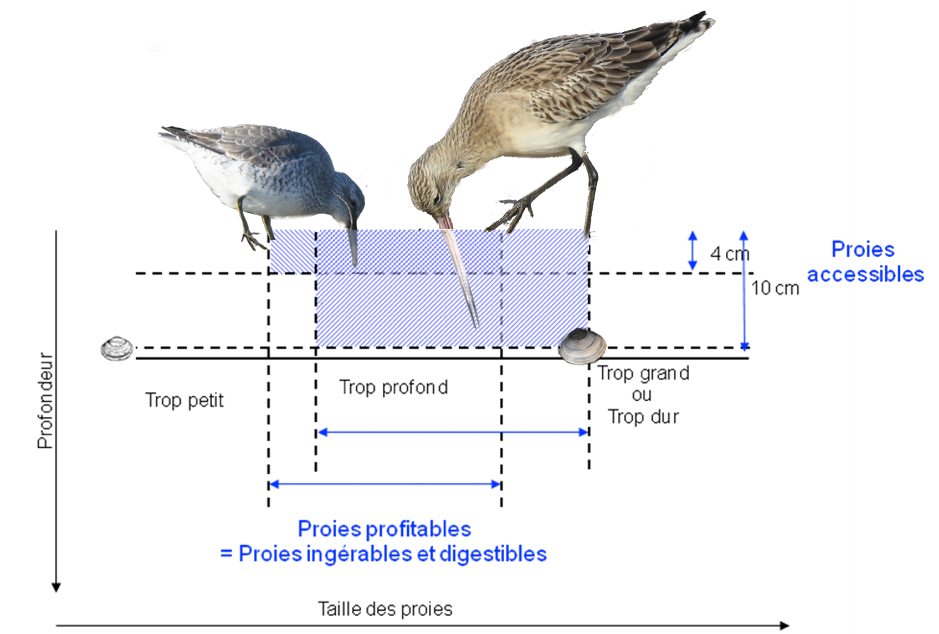
Toutefois, décrire les espèces macrobenthiques présentes sur une vasière, ainsi que leurs abondances et leurs distributions, ne signifie en rien que l’on soit en mesure de définir le stock réel de proies « collectables » par les limicoles. En réalité, sur chaque vasière, seule une fraction de la biomasse totale de la macrofaune est accessible et/ou profitable aux oiseaux, et ce en fonction des caractéristiques morphologiques et physiologiques propres à chacune des espèces. On peut définir cette fraction comme celle répondant à la fois à des critères de disponibilité et de profitabilité. Les critères retenus pour les proies dans ce cadre sont (1) l’accessibilité (2) la détectabilité et (3) l’ingestibilité :
- L’accessibilité est déterminée par la longueur du bec des individus. Les oiseaux peuvent uniquement capturer les proies enfouies dans le sédiment qui sont à portée de leur bec. Les tailles des becs varient de 13 mm pour un grand Gravelot Charadrius hiaticula à 15 cm pour un Courlis cendré Numenius arquata. Certaines espèces munies d’un bec relativement court ont développé une certaine acuité qui leur permet de chasser leur proie à vue en surface des vasières comme le Pluvier argenté Pluvialis squatarola.
- La détectabilité des proies varie selon le type de proie et les techniques de capture employées par les oiseaux. La plupart des limicoles détecte leurs proies par le toucher en sondant le sédiment avec leur bec. Le taux de contact est fonction de la surface corporelle des proies et de la surface du bec de l’oiseau ainsi que de la taille de la zone de recherche et de l’occurrence de la proie. Certaines espèces comme le Bécasseau maubèche sont capables de localiser la présence des bivalves par triangulation suite à la réception des ondes transmises et ricochant sur les coquilles à travers l’eau interstitielle du sédiment.
- L’ingestibilité (et digestibilté) est déterminée par la fraction des tailles de proie qui est ingérable et digérable par chacune des espèces. Le calcul de cette fraction s’applique plus particulièrement aux limicoles qui ingurgitent des mollusques en entier. Les limites sont alors régies d’un côté par la taille, la circonférence et par la dureté de la coquille pour les proies et par la taille et la robustesse du tractus digestif des oiseaux spécialisés sur ce type de proies. Les bécasseaux maubèches ont par exemple, un gésier plus musculeux et proportionnellement plus gros que les autres espèces de limicoles.
Par conséquent, nous sommes tenus de prendre en compte ces trois critères si l’on veut établir précisément la qualité et le potentiel trophique de chacun des sites. La difficulté, en l’occurrence, consiste à isoler la fraction de proies accessibles et profitables du stock total de macrofaune benthique. Pour cela nous devons déterminer (1) la proportion de proies dont la profondeur d’enfouissement reste à portée des espèces étudiées, (2) la proportion de proies détectables par les prédateurs en fonction des techniques et (3) l’intervalle de classe de tailles des proies qui demeure profitable en fonction des taux d’ingestion et dont la limite supérieure correspond à la taille maximale ingérable. Le deuxième critère reste très difficile à évaluer et le plus souvent seuls les deux autres sont pris en compte pour l’évaluation du stock réel de proies
Comment décrire la fraction « collectable » ?
Concrètement la proportion de proies accessibles aux oiseaux est établie à partir d’un fractionnement des carottes prélevées. La partie supérieure des carottes est découpée à la profondeur correspondant à la taille moyenne des espèces étudiées. Cette profondeur de sédiment correspond à 2 cm pour le Tadorne de Belon, à 4 cm pour le Bécasseau maubèche et à 10 cm pour la Barge à queue noire. Les deux portions de carotte sont tamisées séparément. Ne sont retenus que les individus situés à la profondeur accessible et ayant une taille profitable et ingérable. La gamme de tailles profitables est, elle, établie à partir des tailles des coquilles reconstituées en fonction des fragments retrouvés dans les fèces. Cette reconstitution des tailles favorables reste plus difficile pour les annélides, qui offrent très peu d’éléments persistants dans les fèces. Cet exercice a été répété pour chacune des espèces étudiées et pour chacun des sites prospectés.
En ce qui concerne la Barge à queue noire nous avons montré qu’une faible part des organismes était localisée au delà des 10cm de profondeur (Thèse F. Robin, Chap. 4). En conséquence les limicoles à long bec comme les barges ou les courlis sont plus restreints par la taille des proies que par leurs accessibilités dans le sédiment bien que le temps pour extirper des proies plus profondes nécessite un temps de manipulation plus coûteuse en énergie.
Les proies en migration post-nuptiale
Le suivi de la macrofaune benthique n’a pas été limité à la période hivernale puisqu’un même travail a été entrepris au cours des migrations printanières du Bécasseau maubèche par des prélèvements ponctuels directement sur les zones d’alimentation au cours des années 2005 et 2006 et par trois quadrillages au cours de l’année 2006. Cette étude a permis de mettre en évidence les variations inter-saisonnières de la qualité des proies avec des espèces telles que Macoma balthica qui ont une qualité nutritive nettement marquée du fait de l’augmentation de leur masse par la production des gonades (Beukema et al. 1993, Piersma et al. 1994).
Les variations temporelles
Si les suivis en parallèle de plusieurs sites nous informent sur les variabilités spatiales des ressources trophiques susceptibles d’expliquer les distributions des oiseaux, ils ne permettent évidement pas d’expliquer les variations d’effectifs selon une composante temporelle, qu’elles soient d’ordre saisonnier ou annuel. L’objectif serait alors de déterminer si les fluctuations annuelles des effectifs d’oiseaux en hivernage sur les vasières sont corrélées aux fluctuations des ressources trophiques à la même échelle. Les effectifs n’étant également pas constants sur chaque site durant la période hivernale, il est essentiel de tester en premier lieu si les variations d’effectifs peuvent expliquer les variations d’abondance de la ressource au cours des mois d’hiver. Ce travail d’analyse temporelle a été initié dans le cadre de la thèse d’Anne Philippe (2013-2016)
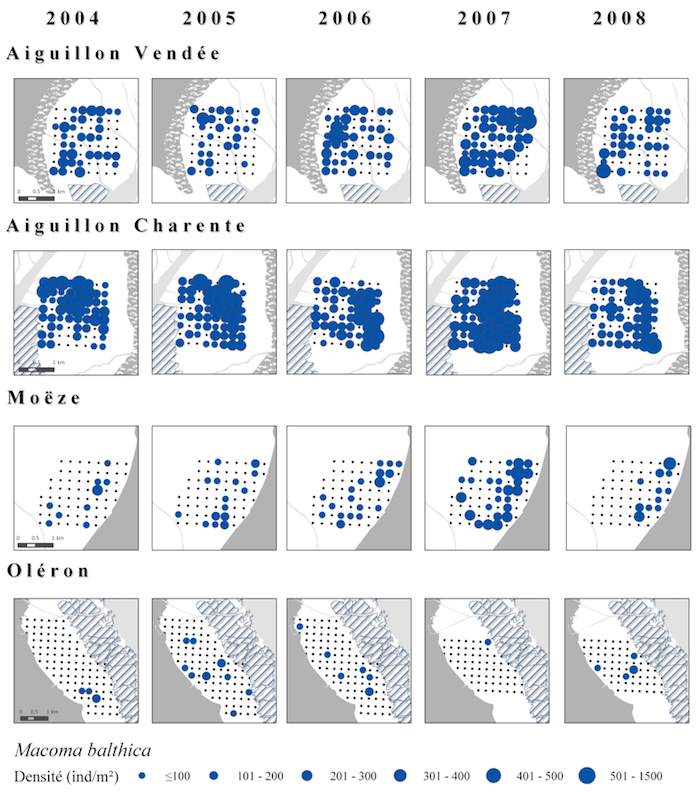
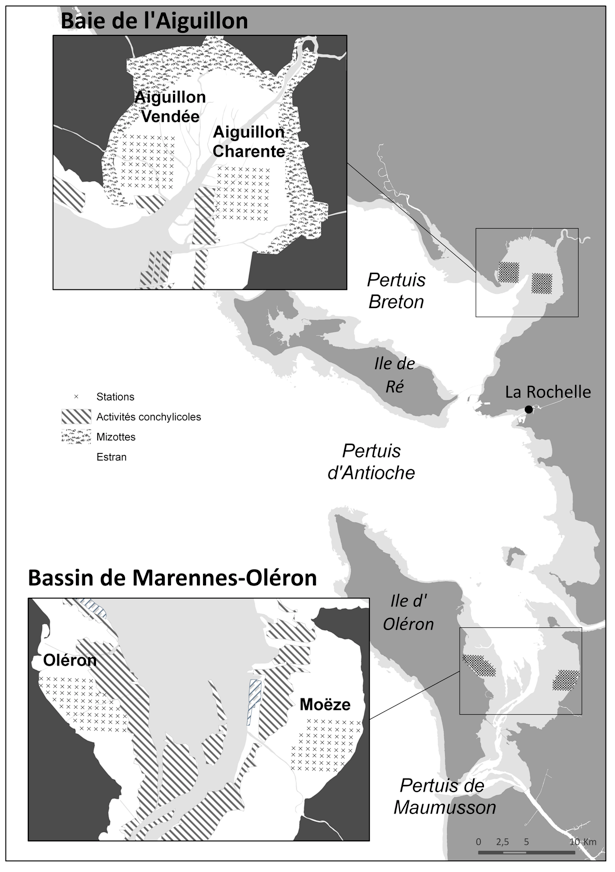
Dans cette optique, et ce depuis la première année de prélèvement en 2004, un suivi annuel à moyen terme des variations d’abondance et de distributions des principales espèces de la macrofaune benthique a été mis en place. Il s’agit d’un suivi sur quatre secteurs, deux en Baie de l’Aiguillon et deux dans le Bassin de Marennes-Oléron couverts selon un même quadrillage comportant 64 stations chacun (Fig. X). Les prélèvements sont effectués au cours des mois de janvier et février au pic de présence maximale des limicoles et durant une période où les dynamiques et les biomasses des populations d’invertébrés sont les plus stables.
Les données de ce « monitoring » de la ressource trophique sont regroupées dans une base de données dont la description à fait l’objet d’un article expliquant Sa structure et son contenu.
Il s’agira d’établir s’il existe des variations spatio-temporelles de ces ressources et si celles-ci peuvent influer sur les capacités d’accueil des sites pour les oiseaux. Une première analyse a été faite en ce sens à partir des six premières années du suivi sur le Bécasseau maubèche (Thèse G. Quaintenne Chap. 5). Cette étude préliminaire montre déjà des variations dans les distributions et densités pour certaines espèces comme M. balthica comme illustré sur la figure suivante :